





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
核糖核酸
与DNA的比较
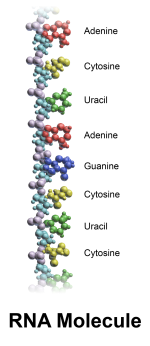
RNA分子中的含氮碱基
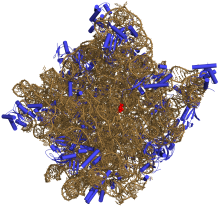
50S核糖体亚单位的3D结构示意图。rRNA以土黄色表示,蛋白质以蓝色表示。活性位点为rRNA的一个小片段,以红色标出
RNA的分子结构与DNA非常相似,但是,两者有以下三个主要不同点:
与DNA相似,大部分有生物活性的RNA,包括mRNA、tRNA、rRNA、snRNA,以及其他一些非编码RNA,虽然是单链,但含有自我互补的序列,能使得它们能进行折叠 ,形成互补双链接构(茎)。对RNA的分析表明,它们有着相对更复杂的结构。和DNA不同,RNA的二级结构并不是单纯的双螺旋,而由一系列短的二级结构构成。通过这些短的二级结构的组合,RNA甚至可以拥有与蛋白质相似的结构,并像酶那样催化化学反应(这样的RNA被称为核酶) 。比如,对核糖体进行分析表明,其催化成肽反应的活性位点完全由RNA构成 。
结构
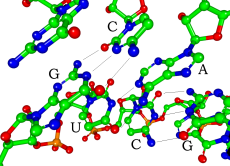
siRNA(小干扰RNA)中的碱基互补配对(图中省略了氢原子)
RNA的单体为核糖核苷酸,其中的戊糖为核糖,依系统命名法可将其中的碳原子从1"编号至5"。含氮碱基与1"碳原子相连。RNA中最基本的四种碱基分别为A(腺嘌呤)、U(尿嘧啶)、G(鸟嘌呤)、C(胞嘧啶)。其中,腺嘌呤和鸟嘌呤为双环的嘌呤,尿嘧啶和胞嘧啶为单环的嘧啶。磷酸基团与一个核糖残基的3"碳原子相连,与下一个核糖核苷酸的5"碳原子相连。磷酸基团在生理pH下,并不都能带上负电荷,因而RNA在生理条件下是带电荷分子(聚阴离子)。C和G、U和A、G和U之间能够形成氢键 。不过,碱基之间也可能发生其他一些相互作用。比如,在一个凸出部分中,一群腺嘌呤可以互相连接 ,GNRA四环中有一个G-A碱基对 。
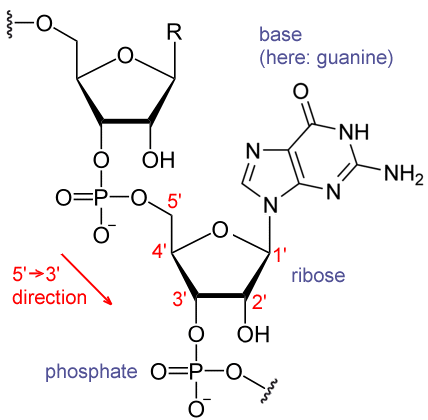
RNA的分子结构
核糖的2位碳上连有羟基为RNA的一个重要结构特点。这类羟基使得RNA双链的结构应与A型构象最接近 ,不过,在单链的某些二核苷酸环境下,也有极小的可能形成DNA最常见的B型螺旋构象 。A型构象使得RNA双链的大沟狭窄而深,小沟浅而宽 。在RNA分子的构象高度可变区域(即不生成双链接构的区域),2"-OH还能攻击附近的磷酸二酯键,使得核糖-磷酸链断裂 。
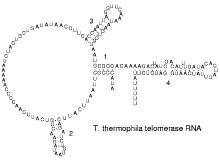
端粒酶RNA的二级结构
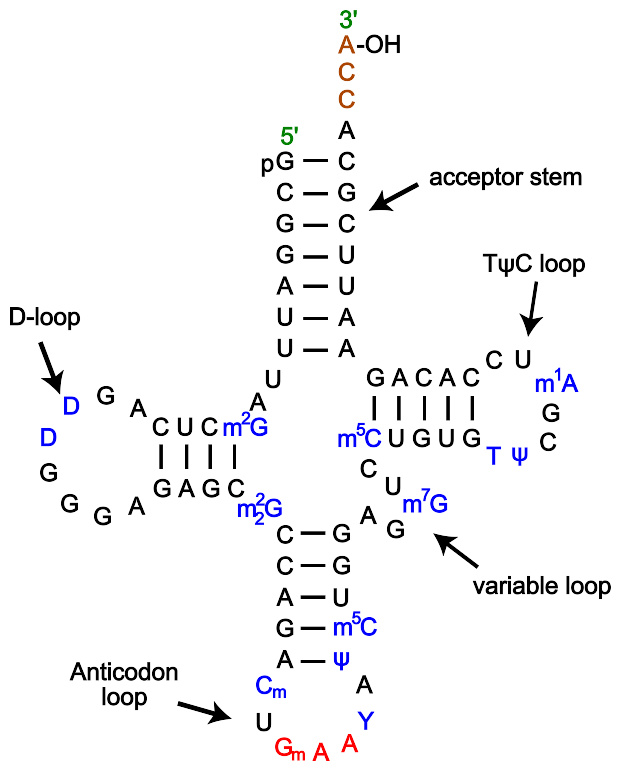
通过转录,仅仅能使RNA链上带A、U、G、C四种含氮碱基 不过,转录后修饰能够通过多种途径对RNA进行改造。比如,转录后修饰能够将稀有碱基 假尿嘧啶 ( 英语 : Pseudouridine ) (Ψ)加到RNA链上。假尿嘧啶与核糖之间的化学键是C-C键而不是尿嘧啶(U)的C-N键。胸腺嘧啶加到RNA链上的情形也很常见(最典型的例子是tRNA的TΨC环) 。另外,次黄嘌呤也是一种常见的稀有碱基。次黄嘌呤为腺嘌呤的脱氨产物,含有次黄嘌呤的核苷被称为肌苷(I)。在基因编码的摆动假说中,肌苷有重要的作用 。
除以上列出的之外,经过编辑的核苷还有100多种 。由修饰引发的结构性变化在tRNA中最为明显 ,假尿嘧啶与经常在rRNA现 2"-甲氧基核糖 ( 英语 : 2"-O-methylation ) 是最常见的修饰产物 。这些修饰的具体作用还没有完全阐明。不过,值得注意的是,在rRNA中,许多的转录后修饰发生在高度功能化的区域,比如肽基转移酶催化中心以及亚基结合部位,似乎说明转录后修饰对RNA发挥正常功能来说相当重要 。
具有催化功能的单链RNA分子,和蛋白质相类似,需要特殊的 RNA三级结构 ( 英语 : Nucleic acid tertiary structure ) 。通过分子内氢键形成的二级结构原件构成了三级结构的框架。二级结构形成了许多可识别的“结构域”——比如茎环结构、膨大结构(bulges)、 内环结构 ( 英语 : internal loop ) 。因为RNA分子带电荷,不少二级结构和三级结构需要Mg 等金属离子来进行稳定 。
自然界中的RNA均是由D-核糖核苷酸聚合而成的D-RNA。使用L-核糖核苷酸则可合成L-RNA。L-RNA对RNA酶的耐受力要强得多 。
合成与修饰
RNA的合成一般由RNA聚合酶催化。RNA聚合酶以DNA为模板,通过转录合成RNA。转录起始于RNA聚合酶与启动子的结合(启动子一般位于基因的上游)。因为RNA聚合酶自带解旋酶活性,仅依靠RNA聚合酶即可实现DNA双链的解开。转录过程中,RNA聚合酶以3"端至5"端的方向读取DNA模板链,并以5"端到3"端的方向合成与之反向平行互补的RNA链。转录的终止由终止子介导。原核生物的终止子有两类:简单终止子与ρ因子依赖性终止子。简单终止子仅靠RNA形成二级结构即可终止转录,而后者在ρ因子的作用下才可以使转录终止。真核生物的转录终止则与转录后修饰密切相关 。
在真核生物中,RNA的 初始转录物 ( 英语 : Primary transcript ) 通常会经过转录后修饰。比如,真核生物的mRNA大都会被加上Poly(A)尾(多腺嘌呤尾巴)以及5"端帽,mRNA前体中含有的内含子序列也会被剪接体切除 。
一些RNA是由RNA复制酶(RNA依赖性RNA聚合酶)以RNA为模板催化合成的。比方说,RNA病毒通过RNA复制酶复制其遗传物质 。另外,RNA复制酶亦参与了众多生物体的RNA干涉过程 。
分类
在细胞中,根据结构功能的不同,RNA主要分三类,即tRNA、rRNA,以及mRNA。mRNA是依据DNA序列转录而成的蛋白质合成模板;tRNA是mRNA上遗传密码的识别者和氨基酸的转运者;rRNA是组成核糖体的部分,而核糖体是蛋白质合成的机械。
细胞中还有许多种类和功能不一的小型RNA,像是组成剪接体(spliceosome)的snRNA,负责rRNA成型的snoRNA,以及参与RNAi作用的miRNA与siRNA等,可调节基因表达。而其他如I、II型内含子、RNase P、HDV、核糖体RNA等等都有催化生化反应过程的活性,即具有酶的活性,这类RNA被称为核酶。
调控RNA
许多种类的RNA,能够透过与mRNA或DNA上的基因片段,部分互补的方式,来调降基因表达。例如在真核生物细胞内,所发现的微RNA(miRNA; 21-22 nt),能引发RNA干扰。miRNA与酵素复合体,会切碎mRNA,阻止该mRNA被翻译,或加速其降解。
虽然小干扰RNA(siRNA; 20-25 nt)的产生,通常是由分解病毒RNA得到,然而也存在内源性的siRNA。 而siRNA引发RNA干扰的机制类似miRNA,有些miRNA和siRNA,能造成其目标基因被甲基化,从而促进或抑制该基因的转录。. 此外,在动物生殖细胞内,所活跃的Piwi-interacting RNA(piRNA; 29-30 nt),被认为能预防转座子,并在配子的发生上,扮演重要角色。
许多的原核生物,具有CRISPR RNAs,其作用机制类似于真核生物的RNA干扰。 其中反义RNA(Antisense RNAs)是最常见的,大多数能调降基因表达,但也有少部分会激活转录进行。 反义RNA的作用机制之一,是借由与mRNA互补配对,来形成双股RNA,而被酵素降解。 此外,在真核细胞内,也许多能调控基因的非编码RNA, 一个常见的例子是Xist,它会附在雌性哺乳动物的其中一个X染色体上,造成其去激活。
一段mRNA自身可能带有调控元件,例如riboswitches,在其五端非翻译区(5" untranslated region)或三端非翻译区(3" untranslated region),包含有顺式作用元件(cis-regulatory elements)能够调控该mRNA的活性。 此外,非编码区上也有可能带有,能调控其它基因的调控元件。
修饰其它RNA
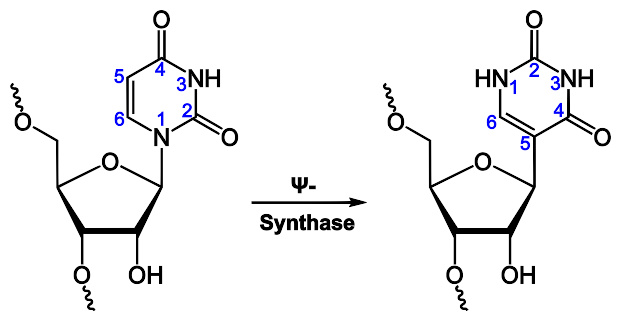
一种的常见RNA修饰,尿苷(Uridine)被转换成假尿苷(Pseudouridine)。
许多的RNA会帮助修饰其它RNA。如前信使RNA(pre-mRNA)中的内含子,会被含有许多核小RNA(snRNA)的剪接体剪接。 或者RNA本身能作为核酶,剪接自己的内含子。
RNA上的核苷酸也可能被修饰,变成非A、U、G、C的核苷酸。在真核细胞中,RNA上核苷酸的修饰,通常是由在细胞核与卡哈尔体中发现的,小核仁RNA(snoRNA; 60-300 nt)所主导。 snoRNA会连结酵素,并以碱基对的方式,引导它们去接上RNA,之后酵素便开始RNA核苷酸的修饰。碱基修饰广泛发生于rRNA与tRNA中,然而snRNA与mRNA也有可能是碱基修饰的目标。 此外,RNA也可能被甲基化。
RNA基因组
如同DNA,RNA也可以携带遗传信息。RNA病毒的基因组由RNA组成,可以翻译出多种蛋白质,其中一些负责基因组的复制,而其它的则作为保护构造,在病毒离开宿主细胞后,保护基因组。类病毒是另一种类型的病原体,但它们仅由RNA组成,且该RNA并不会翻译出任何蛋白质,并利用宿主的聚合酶来复制。
逆转录
逆转录病毒借由将RNA逆转录成为DNA,DNA副本再转录为RNA的方式,来复制他们的基因组。逆转录转座子也利用此方法,来复制DNA与RNA,以完成转座。 此外,真核细胞内的端粒酶,也包含一个作为模板的RNA,利用它来延长染色体端粒。
双链RNA
双链RNA(dsRNA)是指具有两个互补链的RNA,与细胞中的DNA结构相似,它也是某些病毒(双链RNA病毒)的遗传物质。双链RNA如病毒RNA或小干扰RNA(siRNA),可以触发真核生物的RNA干扰,以及脊椎动物的干扰素反应。
发现史

罗伯特·威廉·霍利(左侧)及其研究团队合影。
与RNA相关的研究,造就了许多生物学的发现,以及诺贝尔奖。而核酸于1868年由弗雷德里希·米歇尔(Friedrich Miescher)发现,当时他将该物质称作“核素”,因为它是在细胞核中被找到的。 但不久后,科学家也在没有细胞核的原核生物中,也发现了核酸。此外,早在1939年就有人怀疑,RNA在蛋白质合成中所扮演的角色。 塞韦罗·奥乔亚(Severo Ochoa de Albornoz)与阿瑟·科恩伯格(Arthur Kornberg),因为在实验室内发现了,能够合成RNA的酵素,而获得1959年的诺贝尔生理学或医学奖 然而,之后的研究显示,由他们所发现的酵素多核苷酸磷酸化酶 (英语) ,是负责RNA降解,而非RNA合成。
罗伯特·威廉·霍利(Robert W. Holley)于1965年,发现酵母菌里大小为77个核苷酸的tRNA序列, 并于1968年与哈尔·葛宾·科拉纳(Har Gobind Khorana)以及马歇尔·沃伦·尼伦伯格(Marshall Warren Nirenberg)共同获得了诺贝尔生理或医学奖。在1967年,卡尔·乌斯推测RNA可能具有催化能力,并提出建议指出,最早的生命形式(自我复制的分子)可能依赖于RNA,来携带遗传信息和催化生化反应,即RNA世界学说。
逆转录病毒及逆转录酶,于1970年代早期被发现的,使人们了解到RNA能被逆转录为DNA(与中心法则的一般情况,DNA转录为RNA相反)。 这项发现,使戴维·巴尔的摩(David Baltimore)、罗纳托·杜尔贝科(Renato Dulbecco)与霍华德·马丁·特明(Howard Temin),共同获得了1975年的诺贝尔生理学或医学奖。此外在1976年,瓦尔特·菲尔斯 (英语) 以及他的团队,首度确定了RNA病毒完整基因组的碱基序列(噬菌体MS2)。
在1997年,菲利普·夏普(Philip Sharp)与理察·罗伯茨(Richard Roberts),因为发现哺乳类动物病毒及细胞基因中,具有内含子且会发生RNA剪接,而获得1993年的诺贝尔生理学或医学奖。具有催化功能的RNA(核酶)在1980年代早期被发现,而使得托马斯·切赫(Thomas Cech)与西德尼·奥尔特曼(Sidney Altman),获得1989年的诺贝尔化学奖。而1990年所发现在碧冬茄属上,导入基因会静默植物体自身相似的基因的现象,现今被认为是RNA干扰的结果。
且大约在同时,大小约22个核苷酸的RNA(现在被称为微RNA),被发现在线虫的发育上,扮演重要角色。 而于RNA干扰的研究,让安德鲁·法厄(Andrew Fire)与克雷格·梅洛(Craig Mello),获得了2006年的诺贝尔生理学或医学奖;而同年的诺贝尔化学奖,得奖原因也与RNA相关(在RNA转录上的研究),由罗杰·科恩伯格(Roger Kornberg)获得。此外调控RNA的发现,促使了RNA药物的开发,如利用小干扰RNA来静默目标基因。
参见
脱氧核糖核酸(DNA)
核苷酸
核酸
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读
知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}