





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
糖酵解
糖酵解作用的总反应式
糖酵解作用的总体反应式:
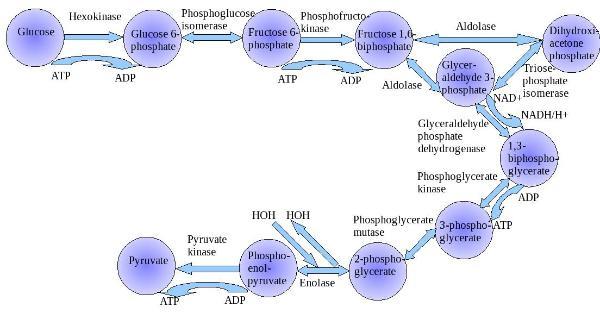
糖酵解作用
对于简单发酵,一个葡萄糖分子到两个丙酮酸分子的代谢具有净产率2个ATP分子。然后,大部分细胞将进行进一步的反应,以“偿还”用过的NAD .,并产生最终产物乙醇或乳酸。许多细菌使用无机化合物作为氢的受体来再生NAD 。
细胞进行有氧呼吸合成更多的ATP,但不作为糖酵解作用的一部分。这些进一步的有氧反应使用来自糖酵解作用的丙酮酸和NADH + H 。真核生物有氧呼吸从每个葡萄糖分子产生额外的约34个ATP分子,但是其中大部分是由截然不同的机制,以在糖酵解的底物水平磷酸化产生的。
糖酵解作用的场所
糖酵解作用在细胞的细胞质中进行。早先人们只知道糖在无氧环境下可降解为乳酸,但今天人们终于清楚知道,不论有氧还是无氧环境,糖会经过同样的过程分解为丙酮酸。不同的则是在有氧条件下,丙酮酸被移出一分子的二氧化碳,剩余的二碳以不稳定的键结连接至辅酶A(一种衍生自维生素B 5 的含硫化合物),形成具有异常活泼乙酰基(附着的乙酸盐)的化学修饰物乙酰辅酶A,从而进入三羧酸循环。
在原核生物和真核生物的大部分缺氧细胞或组织(骨骼肌)中,丙酮酸会转化成乳酸,或者像糖类被酵母分解那样,转化为乙醇和二氧化碳(CO 2 )。在有氧环境下工作的组织(典型:心肌细胞)分解三碳的丙酮酸为乙酰辅酶A和二氧化碳,乙酰辅酶A会进一步行三羧酸循环分解为CO 2 和氢。氢会与氢载体烟酰胺腺嘌呤二核苷酸(NAD )和黄素腺嘌呤二核苷酸(FAD)结合成(NADH和FADH 2 )。在线粒体里进行的呼吸链,NADH和FADH的氧化会导致ATP的产生,能量会储存在ATP的高能磷酸键供细胞使用。
糖酵解作用是唯一一条现代生物都具有的代谢途径,出现时间很早。糖酵解作用最早可能发生在35亿年前第一个原核生物中。
糖酵解步骤
糖酵解的第一步是葡萄糖磷酸化为6-磷酸葡萄糖。不同细胞类型中所含有的酶也不一样,在所有的细胞中,皆有己糖激酶进行催化,而在肝和胰腺中,则另外含有一种称为葡(萄)糖激酶(己糖激酶IV)的酵素 。磷酸化过程消耗一分子ATP,后面的过程证明,这是回报很丰厚的投资。细胞膜对葡萄糖通透,但对磷酸化产物6-磷酸葡萄糖不通透,后者在细胞内积聚并继续反应,将反应平衡向有利于葡萄糖吸收的那一面推移。之后6-磷酸葡萄糖会在磷酸己糖异构酶的催化下生成6-磷酸果糖。(在此果糖也可通过磷酸化进入糖酵解作用途径)
接着6-磷酸果糖会在磷酸果糖激酶的作用下被一分子ATP磷酸化生成1,6-二磷酸果糖,ATP则变为ADP。这里的能量消耗是值得的,:首先此步反应使得糖酵解作用不可逆地继续进行下去,另外,两个磷酸基团可以进一步在醛缩酶的参与下分解为磷酸二羟丙酮和3-磷酸甘油醛。磷酸二羟丙酮会在磷酸丙糖异构酶帮助下转化为3-磷酸甘油醛。两分子3-磷酸甘油醛会被NAD +和3-磷酸甘油醛脱氢酶(GAPDH)的氧化下生成1,3-二磷酸甘油酸。
下一步反应,1,3-二磷酸甘油酸转变为3-磷酸甘油酸。此反应由磷酸甘油酸激酶催化,高能磷酸键由1,3-二磷酸甘油酸转移到ADP上,生成两分子ATP。在此,糖酵解作用能量盈亏平衡。两分子ATP消耗了又重新生成。ATP的合成需要ADP作原料。如果细胞内ATP多(ADP则会少),反应会在此步暂停,直到有足够的ADP。这种反馈调节很重要,因为ATP就是不被使用,也会很快分解。反馈调节避免生产过量的ATP,节省了能量。磷酸甘油酸变位酶推动3-磷酸甘油酸生成2-磷酸甘油酸,最终成为磷酸烯醇式丙酮酸。磷酸烯醇式丙酮酸是高能化合物。最后,在丙酮酸激酶的作用下磷酸烯醇式丙酮酸生成一分子ATP和丙酮酸。此步反应也受ADP调节。
反应序列
糖酵解作用反应序列可被分为十个步骤
准备阶段
通常视前五步为准备(或投入)阶段,因为这些步骤消耗能量以将葡萄糖转变为两个丙糖磷酸,即甘油醛-3-磷酸和磷酸二羟丙酮。
放能阶段
糖酵解作用的第二阶段为放能阶段,此阶段的目的在于产生高能分子ATP和NADH。因为一个葡萄糖在准备阶段时已经变成两个丙糖,所以在放能阶段中每个反应会发生两次。最后产生2个NADH和4个ATP,使得单一葡萄糖在经过整个糖酵解作用后净得2个NADH和2个ATP。ATP会用于其他需能反应,而NADH则会进入呼吸链或作为还原剂参与细胞内其他还原加氢反应。
糖酵解中的不可逆反应
人体可通过糖异生,即从非糖化合物,如丙酮酸和乳酸等物质重新合成葡萄糖。当肝或肾以丙酮酸为原料进行糖异生时,糖异生中的其中七步反应是糖酵解中的逆反应,它们有相同的酶催化。但是糖酵解中有三步反应,是不可逆反应。在糖异生时必须绕过这三步反应,代价是更多的能量消耗。
这三步反应都是强放能反应,它们分别是:
1葡萄糖经已糖激酶催化生成6磷酸葡萄糖ΔG= -33.5 kJ/mol
2 6磷酸果糖经磷酸果糖激酶催化生成1,6二磷酸果糖ΔG= -22.2 kJ/mol
3磷酸烯醇式丙酮酸经丙酮酸激酶生成丙酮酸ΔG= -16.7 kJ/mol
糖酵解作用中的调节位点
糖酵解作用在体内可被精确调节,这样一方面可以满足机体对能量的需要,另一方面又不会造成浪费。同时,当细胞内还进行糖异生的时候,调节就显得非常重要了,因为要避免空循环的发生。
调节是通过改变底物浓度,酶的活性实现的。
磷酸果糖激酶是其中最重要的限速酶,这也是巴斯德效应的关键参与者,它也决定了糖异生的速度,成为调节位点。AMP的浓度越高,酶的活性越高。就是当机体大量消耗了ATP,而相应又产生了很多AMP的时候,酶的活性提高,使得糖酵解按生成ATP的方向快速前进,以提高ATP产量。
NADH的去路
在细胞中,NADH与NAD 是处于动态平衡的。在糖酵解过程中生成的NADH必须被进一步氧化,转化为NAD 才能够让糖酵解持续进行。另外足够的NAD 是3磷酸甘油醛成为1,3二磷酸甘油酸这一步反应重要的前提。在此过程中NAD 会被还原为NADH+H ,即是氢载体,通过穿梭将氢带到呼吸链。
NAD 的再生可通过这三种不同的过程来实现。
乳酸脱氢酶:由丙酮酸形成乳酸,此过程发生在骨骼肌及部分微生物中。
乙醇脱氢酶:经丙酮酸脱羧酶将丙酮酸转变为乙醛,再由乙醛经乙醇脱氢酶催化还原形成乙醇,此过程发生在大多数植物和一些产乙醇的微生物中。人类基因组可以编码乙醇脱氢酶,但是并没有编码丙酮酸脱羧酶的基因,因此无法将丙酮酸转化为乙醇。
线粒体穿梭:经线粒体穿梭途径进入线粒体的呼吸链生成ATP。
能量转化
平衡点
值得一提的是,生成1,6-二磷酸果糖后的大部分反应都是向能量升高的方向进行的,没有酶(磷酸果糖激酶( PFK ),磷酸甘油酸激酶( PGK ))的催化,是不会自发进行的。而糖酵解的逆过程--糖异生(从甘油等非糖物质生成葡萄糖)则容易进行,此过程用到大部分在糖酵解里面出现过的酶,除了提到的两位“车夫”外,它们只出现在糖酵解中。在糖异生这两步逆反应会放出大量的热,分别为-14及-24 kJ/mol。
无氧环境和有氧环境的能量转化
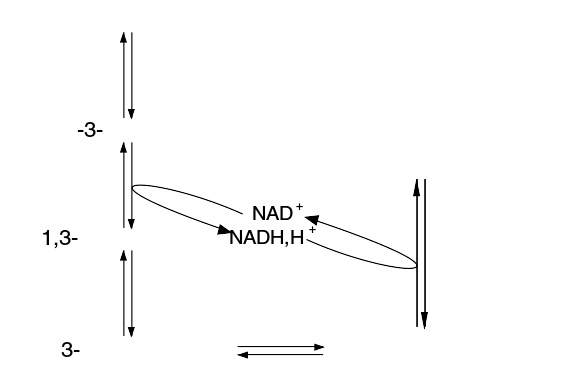
糖酵解中的NAD+和NADH+H 循环
在糖酵解作用中,每分子葡萄糖提供两分子ATP。真核生物的粒线体能同时从两分子丙酮酸中另外获得36分子ATP。能量转化的多少取决于在细胞质中产生的NADH + H 通过粒线体膜的方式。
不论在无氧还是有氧环境中,糖酵解成丙酮酸这一过程都能进行。3-磷酸甘油醛在3-磷酸甘油醛脱氢酶GAPDH的作用下脱氢。脱下的氢离子会将氧化剂(辅酶)NAD 还原成NADH + H 。NAD 会在呼吸链中再生。若在无氧环境,放热的(ΔG ´ = - 25 kJ/mol)乳糖脱氢酶(LDH)反应会再生NAD :丙酮酸的还原会生成乳糖和再生NAD (酵母则会使用另外两种酶—丙酮酸脱羧酶加乙醇脱氢酶)。下图可阐明此过程:
无氧环境下糖酵解作用GAPDH-和LDH-反应的相互联系,除了少部分NADH+H 会被磷酸甘油脱氢酶(GDH)转化外,大部分会用于再生NAD 。
参看
三羧酸循环
发酵
糖异生
磷酸戊糖途径
发酵
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。




- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}